一般名:horned passalus科学名:Odontotaenius disjunctus(Illiger)(Insecta:甲虫目:Passalidae:Passalinae)
主に新世界熱帯地域で500種近くのPassalidaeが記載されている(Arnett et al. 2002). 一般的にベスバグまたはパテントレザーカブトムシとして知られている家族Passalidaeは、スーパーファミリー Scarabaeoideaのメンバーであり、米国内で発生する種はわずかである(Schuster1983)。, 角のあるpassalusまたはbetsy beetle、Odontotaenius disjunctus(Illiger)は、広く分布し、容易に認識できるカブトムシであり、北アメリカの地理における相対的な独占のために、米国で最も一般的に遭遇するPassalidaeのカブトムシである。 以前は、アメリカではPassalus punctiger(Lepeletier)とPassalus punctatostriatus(Percheron)が外来種として報告されていましたが、最近の記録ではどちらの種の現在の個体群も示されていません(Schuster1983)。, 角のあるpassalusを含む家族Passalidae内の種は、木材の有益な分解剤です。 角のあるpassalusは腐敗した木材や丸太を分解するだけであり、都市構造の害虫ではありません。
図1. 角のあるパッサルス、Odontotaenius disjunctus Illigerの側面図。 光沢のある黒い色は、別の一般的に使用される名前を担当していた:パテントレザービートル。 フロリダ大学のライル-J-バスによる写真。,

Odontotaenius floridanus Schuster、O.disjunctusに近い祖先のカブトムシは、フロリダ州の限られた地域で固有に発生します。 Odontotaenius floridanusは、より広い前tib骨と減少した角を有することを除いて、ライフサイクルおよび形態のほとんどの面でO.disjunctusに似ている(Schuster1994)。 アメリカ合衆国ではPassalidaeのメンバーはほとんどなく、以前に言及された二つの種のみが中米から移住または導入されたとは考えられていない(Schuster1983)。,
シノニム(Back to Top)
Odontotaenius disjunctusは、以前はPopilius disjunctus(Illiger)およびPassalus cornutus(Fabricius)(Hincks1951)としても知られていた。
Horned passalusはアメリカ昆虫学会の承認された一般的な名前ですが、次の一般的な名前もしばしばそれに関連付けられています:betsy beetle、bess bug、patent leather beetle、Jerusalem beetle、horn beetle、peg beetle。 これらの一般的な名前のいくつかは、O.disjunctusに特に適用され、他の人は一般的に家族Passalidaeを参照することができます。,
分布(トップに戻る)
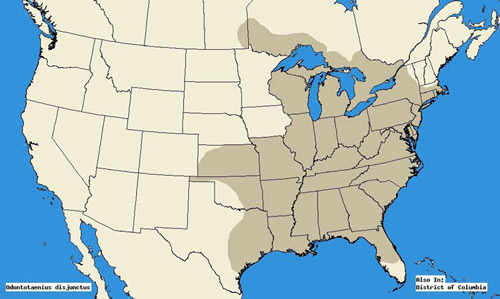
角のあるpassalusは、フロリダ州中部からマサチューセッツ州、テキサス州南部からミネソタ州、ネブラスカ州までの広い範囲で発生します(Schuster1983)。 より最近の情報源は、テキサス州東部から、米国東部全体、およびマニトバ州南部からカナダの落葉樹林、およびオンタリオ州南部までのさらなる拡大を示している(Arnett et al. 2002).
図2. オドントタエニウス-ディジュンクトゥス-イリゲル(Odontotaenius disjunctus Illiger)は、アメリカ合衆国の小惑星。, (地図は、文献ソースSchuster1983およびArnett et al. 2002). マイク-ブーン、クリストファー-S-ビブス、フロリダ大学によってプログラム。
Description(Back to Top)
大人:角のあるpassalus大人は比較的大きく、暗い光沢のある黒いカブトムシであり、大人のサイズは通常30–40mm(1.2inch-1.6inch)の範囲である。 金色の毛は、脚、前脚、触角の中央のペアを裏打ちするのを見ることができます。 これらの毛は肉眼によって探索可能です。
図3., 角のあるパッサルス、Odontotaenius disjunctus Illigerに金色の髪の縞があります。 クリストファー-S-ビブス、フロリダ大学による写真。

elytra、または硬化した外側の前縫いは、深い溝によって特徴付けられる。 角のあるpassalusはまた、pronotumの正中線の下に溝を持っています。
図4. 大人の角のあるpassalus、Odontotaenius disjunctus Illigerの背面図。 フロリダ大学のライル-J-バスによる写真。,

アンテナには10個のセグメントがあり、遠位の三つのセグメントは薄板クラブを形成しています。 これは家族の容易に識別可能な特徴である。
図5. ラネラテアンテナ、角のあるpassalus、Odontotaenius disjunctus Illigerの特徴のクローズアップ。 クリストファー-S-ビブス、フロリダ大学による写真。

単一の湾曲したホーンは、頭の上と目の間を前方に指します。, 角のあるpassalusの大人はさらにはっきりとまた目に見える厚く、強いあごを持っています。
図6. 頭と角のあるpassalus、Odontotaenius disjunctus Illigerの湾曲した角のクローズアップ。 また、金色の毛の縞と頭のすぐ後ろのダニに注意してください。 フロリダ大学のライル-J-バスによる写真。

平均して、成人は一年まで生きています。, 角状のpassalusの大人の主特徴は妨げられるか、またはかき混ぜられたとき出す音響信号(ぜん鳴)である。
卵:角のあるpassalusの卵は非常に大きく、最初に敷設されたときに3.0×2.4mm(約0.1インチ)で測定し、孵化する直前に3.7×3.2mmに達します。 卵の大きさは、大人の大きな下顎骨で操作することが容易になります。 卵は通常、成虫からのフラスに囲まれたグループで見つかります。
幼虫:幼虫は三つのインスターまで持っている大きな、白い草です。, 幼虫は成虫と同じギャラリーで見つかります。 栄養素を得るために、幼虫は成虫から消化された木材を食べる。
図7. 角のあるパッサルスの幼虫,Odontotaeniusdisjunctusinligerは体の形を見ることができた。 フロリダ大学のライル-J-バスによる写真。
図8. 角のあるpassalusの幼虫、Odontotaenius disjunctus Illiger、頭のカプセルのクローズアップ。 フロリダ大学のライル-J-バスによる写真。,
図9. 角のあるpassalusの幼虫、Odontotaenius disjunctus Illiger、脚のクローズアップ。 フロリダ大学のライル-J-バスによる写真。



グラブは足の二つのペアしか持っていないように見えますが、第三のペアが存在し、減少しています。 最初の二つのペアのみが移動に使用されます。 Scarabaeoidaeスーパーファミリー内のカブトムシのために典型的であるように、grubsはアクティブでないときに特徴的なC字型の姿勢を保持します。, 幼虫はよく比例した下顎を除いて積極的な防御を持たず、成体のカブトムシによって使用されるものと同様の聴覚シグナルを持っていません。 P>
蛹:蛹が形成され始めると、彼らは虹の光沢を持つ真珠のような白になります。 蛹の年齢として、彼らは彼らの虹の光沢を失い、オフホワイトからアーストーンまでの色の範囲であることができます。 出現に近づいている蛹は半透明になります。 蛹化時間は気候条件によって異なります。
図10. 角のあるパッサルスの蛹、Odontotaenius disjunctus Illiger。, シェリー-コックス、ミズーリ州保全省による写真。

ライフサイクルと生物学(トップに戻る)
その大きな下顎骨を使用して、角のあるpassalusは落ちた丸太に切り取り、それが住んでいると繁殖
図11. 角のあるpassalus、Odontotaenius disjunctus Illigerの深く刻まれた木製のトンネル。 (フロリダ大学教育学部)。 クリストファー-S-ビブス、フロリダ大学による写真。,

大人は、木材が木材の最初の住民よりも多くをサポートするのに十分な大きさであれば、落ちた木材のセクションを集約し、競います。
図12. 同じ木片における角のあるpassalus、Odontotaenius disjunctus Illigerの集合。 (フロリダ大学教育学部)。 クリストファー-S-ビブス、フロリダ大学による写真。,

新しい卵クラッチの親は、両性の成虫がひなの協調的ケアにおいてギャラリー複合施設を通して幼虫を傾向がある亜社会的関係を作り出す(Schuster and Schuster1985)。 木材が邪魔されずに残っている場合、複数の世代が同じ丸太を同居させ、若い大人も兄弟と新しいひなのケアに参加します。 成虫は縄張りであり、侵入者から自分のギャラリーを守るが、成虫と幼虫は共食いする(King and Fashing2007)。, ダニは、一般的に角のあるpassalusの外骨格に見られます。 これらのダニは多様ですが、ヒトやカブトムシには有害ではありません。
卵が産まれると、成体は卵のクラッチを繰り返しギャラリーを通って移動します。 幼虫が定着するための最良の木材の領域を探して、卵の両親は、卵の保護と輸送のための理想的な摂食場所を見つけて守ろうとします(Wicknick and Miskelly2009)。 卵が孵化すると、幼虫は親の成虫と一緒に残り、フラスと軟化した木材の混合物を共有します。, 温度および湿気の状態によって、全体のライフサイクルは夏の季節の内に起こるか、または14-16か月長く取るかもしれない(Krausおよびライアン1953)。
蛹化の前に、成体は腐敗した木材の中に蛹室細胞を残骸と残骸で詰め込みます。 これらの蛹細胞は脆弱な段階を偽装しますが、これらの細胞の完全な目的は証明されていません(Valenzuela-Gonzalez1992)。 若い新しく出現した大人を含むすべての年齢の大人は、これらの蛹細胞を作り出す能力を持っています。 細胞の質は、成人の年齢とともに変化する。, 蛹のまわりのこれらの残骸のセルが傷つくようになれば、大人はそれらを修理します。 完全な発達のために追加の時間が必要な場合、蛹は越冬することができます。
いくつかのログは、カブトムシの複数の世代を維持することができますが、若年成人は、通常、最小限の時間のために親ログ内に滞在します。 同じホスト構造内のカブトムシの重複世代は、それが完全に成熟するのにかかる限り、それだけで残るでしょう。 これらの若い成虫は、種の特徴的な黒い殻を欠いており、代わりに蛹期から出てくると赤い着色をしています。, この赤い色はゆっくりと黒くなり、その時点で昆虫は成熟した大人とみなされます(Schuster and Schuster1985)。
図13. 新たに出現したteneral大人の角のpassalus、Odontotaenius disjunctus Illigerは、すでにダニ(elytraを這う)が生息していました。 シェリー-コックス、ミズーリ州保全省による写真。

この新しく成熟した大人は、親のログを残して、彼らの翼の使用の唯一のインスタンスの一つである結婚飛行に参加します(MacGownおよびMacGown1996)。, 結婚飛行の観測中に,成体はライトトラップや街灯のような光誘引物質に敏感であることが分かった。 結婚式の飛行の終わりに、大人は別の集計を開始するために新しいログを探し出します。
角のあるパッサルスの穏やかな性質と飛ぶことへの抵抗は、大学による生きた標本のデモンストレーションやアウトリーチ教育のための安全で扱いやすい生き物としての採用に貢献している。, 内政上、それは心配の低い維持で、普通カブトムシのために適しているあらゆる自然な区域で得ることができる湿らせた木の一貫した供給だけを
餌の生息地(トップに戻る)
角のあるパッサルスは、オークなどの広葉樹を好む(Quercus spp.)またはニレ(Ulmus spp.そして、落ちて腐敗し始めた丸太の中にのみ存在します。 カブトムシは乾燥条件に敏感であるため、最適な条件には高い水分レベルが含まれます。, カブトムシが木材を消化するためには、木材は天然に存在する真菌や細菌などの微生物叢を持っていなければならず、木材を分解しています。 これらの微生物叢は、木材の消化前に役立ちます。
植物摂食昆虫では、消化されていない遺体は、フラスと呼ばれる細かい粉末状の材料の形で排出される。 角のあるpassalusが丸太の中に住居を取ると、カブトムシは木材パルプを噛んでフラスを追放することによって丸太の分解を促進する。 次いで、真菌および細菌は、それら自身の栄養抽出のためにfrassを標的とする(Arnett et al. 2002)., フラスが真菌および細菌によって十分に再処理されると、角状のパッサルスはその後、成功した消化のために再びフラスおよび木材を消費する。 角のあるpassalusによって生息されているログは、ログの周りと下に散らばっておがくずを持っている傾向があります。
サウンド(トップに戻る)
家族Passalidaeのいくつかのメンバーは、激突する能力を持っています。 昆虫の他のグループはまたコオロギ、katydids、バッタ、蟻および蝉のさまざまな種を含むがこれらに限定されないこの容量を、開発しました。, 昆虫の鳴き声は、二つの体のセグメントまたは手足の間に摩擦を作成することによって音の放出です。 Stridulationはプラスチック櫛を取り、櫛の歯の列の下のあなたの釘の端を摩擦することによって単に示すことができます。 ストリデュレーションは、昆虫のフェロモンおよび光シグナルに特異性が類似した種内の通信の一形態として一般的である。
角のあるpassalusでは、成虫と幼虫の両方が、激突を使用してはっきりと聞こえる音を作り出す能力を持っています。, 角のあるpassalusは、特に大人と幼虫の間で17の発見された音声信号を持っており、そのすべてが未成熟カブトムシの防衛、グループコミュニケーション、および管理 Reyes-Castillo and Jarman(1980)によって表現されているように、このレパートリーは”節足動物で知られている最も精巧な音のコミュニケーションシステムを表しています。”
大人は適応された腹部と翼の構造を使用して聴覚信号を作成します。, 保護エリトラの下に折り畳まれたままである非硬化後肢の翼は、下側に小さな棘の多くの列を持っています。 後肢の翼のこれらの棘のある領域の下の腹部のセグメントには、pars stridensと呼ばれる強化された領域があります。 この構造は肉眼では識別できません。
図14. Elytraは、ピン留めされ乾燥した角のあるpassalus、Odontotaenius disjunctus Illigerの背側の喉と腹部を露出させ、提起した。 クリストファー-S-ビブス、フロリダ大学による写真。,

音を作るために、棘は腹部のこのテクスチャ領域を横切って掻き取られます。 動きの速度とパターンに応じて、異なる周波数のノイズを生成することができます(Reyes-Castillo and Jarman1983)。
未熟な角のあるpassalusでは、足の第三のペアは、実際にぜん鳴を支援するために減少します。 脚の中央のペアは、脚の近位端にパースストリデンス(尾根のセット)を持っています。, 三番目の組の足は、減らされた放射状の動きと、頂点が付いている標準のstridensに達し、騒音を発生させるのに必要な摩擦を作成できる。 幼虫の聴覚シグナルは、成体のシグナルにはあいまいである。
図15. 角のあるpassalus、Odontotaenius disjunctus Illigerの幼虫における足の減少した第三のペアのいずれかのクローズアップ。 フロリダ大学のライル-J-バスによる写真。,
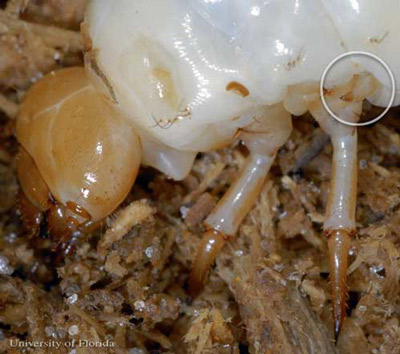
これらの減少した脚の先端には、脚点の頂点に近い複数の歯があります。 これらの様々な歯は、幼虫、親成虫、および非親成虫の間で通信するために必要な異なる周波数を作成するために、異なる角度で使用されます。
角のあるpassalusの鳴き声は、保護ログの外でさえ、防御目的に役立つという証拠を示しています。 幼虫および完全に発達したカブトムシは捕食者に対する防御がほとんどない。 しかし、その用意がある音声が乱れに貢献する。,
Odontotaenius disjunctusによって放出された防御音のオーディオファイルを聴くにはここをクリックしてください。
防御としての音作りの重要性をテストする実験では、角のあるpassalusの幼虫は、捕食を観察するために意図的にカラスにさらされました。 死亡率までの時間の長さは、積極的に音を放出した幼虫の間ではるかに大きかった(Buchler et al. 1981).
選択された参考文献(トップに戻る)
- Arnett Jr RH,Thomas MC,Skelley PE,Frank JH. (編集部)2002年。 アメリカのカブトムシ、ボリュームII:Polyphaga:Curculionoideaを通じてScarabaeoidea。 ペットショップ, 861. CRCプレス、ボカラトン、フロリダ州。
- Buchler ER,Wright TB,Brown ED. 1981. パッサリッドカブトムシOdontotaenius disjunctus(甲虫目:Passalidae)による激突の機能について。 動物の行動29:483-486.
- ヒンクスWD. 1951. “Passalus cornutus Fabricius”(パッサルス-コルヌートゥス-ファブリシウス)についてのメモ。 コレオプテリスト紀要5:12-13。
- キングA、ファッシングN.2007。 亜社会性のカブトムシOdontotaenius disjunctus(Illiger)(甲虫目:Passalidae)における幼児行動。 昆虫の行動のジャーナル20:527-536。
- Kraus JB,Ryan MT. 1953., 角のあるpassalusカブトムシ、Popilius disjunctus(Illiger)の発生学における発達の段階。 アメリカ昆虫学協会の年表46:1-20。
- マックガウンJA,マックガウンMJ. 1996. カブトムシOdontotaenius disjunctus(Illiger)(甲虫目:Passalidae)の結婚飛行の観察。 コレオプテリスト紀要50:201-203。
- Reyes-Castillo P,Jarman M.1983. 成虫のカブトムシ(甲虫目:カブトムシ科)の外乱音:構造的および機能的側面。 アメリカ昆虫学協会の年代記76:6-22。,
- Reyes-Castillo P,Jarman M.1980. ネオトロピカルパッサリダ科(甲翅目:Lamellicornia)における幼虫のぜん鳴に関するいくつかのノート。 コレオプテリスト紀要34:263-270。
- シュスター JC. 1994. オドントタエニウス-フロリダヌス(Odontotaenius floridanus)は、アメリカ合衆国のカブトムシ。 フロリダ昆虫学者77:474-479。
- シュスター JC. 1983. アメリカ合衆国のPassalidae。 コレオプテリスト紀要37:302-305。
- シュスター JC. 1975. 比較行動、音響信号、および新しい世界Passalidae(甲虫目)の生態学。 127ページ 博士論文。, フロリダ大学、ゲインズビル、フロリダ州。
- シュスター JC、シュスターLB。 1985. パッサリドカブトムシ(甲虫目:Passalidae)における社会的行動:協力的なブロードケア。 フロリダ昆虫学者68:266-272。
- バレンズエラ-ゴンザレスJV。 1992. パッサリッドカブトムシ(甲虫:Passalidae)におけるさなぎ細胞構築挙動。 昆虫の行動のジャーナル6:33-41。
- 2009. 非同棲ベスカブトムシ、Odontotaenius disjunctus(Illiger)(甲虫目:Passalidae)の間の行動相互作用。 コレオプテリスト紀要63:108-116。